

La diversità dei virus influenzali si spiega principalmente per due aspetti molto correlati tra di loro. Da un lato i meccanismi evolutivi principali tipici di questa tipologia di virus - la derivazione ed il riordinamento o intercambio genetico (già discussi in un altro articolo di questa sezione) - ; da un altro lato, il comportamento epidemiologico del virus in ogni specie, il quale, come discuteremo più avanti, ha un importante effetto sui meccainismi evolutivi.
Se confrontiamo, in base alla derivazione genetica, i virus presenti nei suini con i virus dell'essere umano, risaltano due aspetti: 1) a partire da un ceppo definito, il virus nell'uomo varia antigenicamente (cambi negli amminoacidi) più veloce rispetto al suino, tuttavia il tasso di mutazione genetica del virus è uguale per entrambe le specie e 2) a livello globale, la diversità del virus dell'Influenza, sia dal punto di vista genetico che antigenico, è molto maggiore nei suini rispetto all'uomo. Nonostante questi due aspetti possano sembrare a priori contraddittori, sono, come vedremo in seguito, perfettamente compatibili.

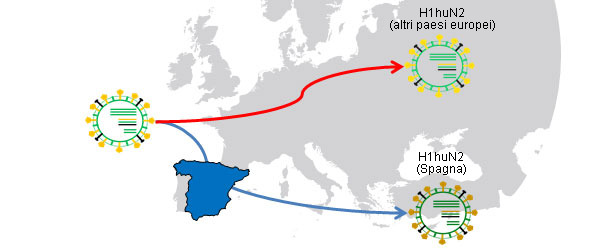
Figura 1. Ancestrali comuni H1huN2 (1994); Isolamento geografico → le derivazioni producono varianti correlate con ogni paese.
In altri articoli di questa sezione, abbiamo visto che l'influenza suina si può comportare in modo endemico o enzootico, detto in altri modi, i virus influenzali possono "resiedere" negli allevamenti. Questa situazione è molto frequente e prevede la comparsa di circolazioni recidive. In un lavoro del 2001 (De Jong e collaboratori) hanno confrontato geneticamente ed antigenicamente oltre 20 tipi di H1N1 suini del "tipo avicolo" (H1avN1). Questi virus erano stati isolati in 6 allevamenti in Olanda che erano endemicamente infetti. In base al gene dell'emagglutinina (H), si è comprovato che gli isolati formavano 5 gruppi che si differenziavano sia a livello genetico sia antigenico. E' interessante sottolineare che ogni gruppo corrispondeva ad un allevamento (eccetto uno che includeva gli isolati di 2 allevamenti. Questo lavoro dimostra che tutti gli isolati hanno un H1avN1 ancestrale comune, ma che grazie alla circolazione endemica, i virus evolvono e si diversificano in forma isolata. In un altro studio (Martín-Valls et al., 2013), realizzato presso il CReSA dal 2009 al 2012, si isolarono 4 virus H1N2. Nell'analizzare l'H1 dal punto di vista genetico, si osservò che tutti appartenevano al tipo H1N2 "tipo umano" (H1huN2), però formavano un gruppo chiaramente diverso dai H1huN2; si potrebbe concludere che vi è stata un'evoluzione isolata in Spagna di questo tipo. Successivamente, abbiamo potuto comprovare che questi virus reagiscono in modo limitato con i sieri che reagivano con un altro H1huN2 commerciale, indicando una certa derivazione antigenica (dati non pubblicati). In definitiva, sia i fattori epidemiologici come l'evoluzione dagli isolati in allevamenti in condizioni endemiche, o l'isolamento geografico, giocano un ruolo importante nella comparsa di nuove varianti antigeniche. Nell'uomo, tuttavia, la situazione è completamente distinta. In questo caso si potrebbe dire che a causa della pressione immunitaria globale, i virus sono costretti a cambiarsi antigenicamente per poter sopravvivere. Oggi le molteplici e svariate connessioni esistenti tra paesi di tutto il mondo eliminano le situazioni di isolamento. Per cui, nell'uomo esiste una seleziona positiva a livello globale delle varianti di virus influenzali in grado di sfuggire alla risposta immunitaria; questo fenomeno si traduce nella necessità costante di aggiornamento dei vaccini nel settore umano.
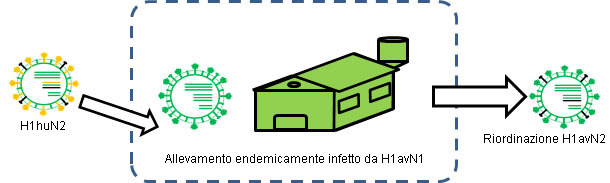
Figura 2. H1huN2 si transmitte ad un'allevamento endemicamente infetto da H1avN1 → Si produce un riordinamento.
Il riordinamento o interscambio genetico può anche essere origine di cambi a livelli antigenici, addirittura in modo immediato. Nell'essere umano, per esempio, questa è stata la causa principale delle ultime 3 pandemie, dato che ha permesso l'introduzione di almeno una nuova emaglutinina diversa antigenicamente alla pre-esistente nella popolazione e di fronte alla quale non vi era immunità precedente. In un altro articolo di questa sezione abbiamo già visto come nel suino i virus influenzali subiscano frequenti riordinamenti, generando di conseguenza nuovi ceppi. Esempio di questo fatto sono l'H1avN2, un H1N2 che si incrocia geneticamente con i virus H1avN1 (Rose et al., 2013), o i H1huN1, che si incrociano antigenicamente con l'H1huN2 (Martín-Valls et al., 2013; Rose et al., 2013). Affinchè questi riordinamenti avvengano è necessario che due ceppi distinti infettino lo stesso soggetto allo stesso tempo. Ovviamente, le situazioni endemiche sono scenari ideali affinchè questo succeda. Infatti, quando le situazioni endemiche sono state valutate, si osservarono le prove di riordinamento genetico tra ceppi diversi, inclusi i subtipi (Martín-Valls et al., 2013; Rose et al., 2013).
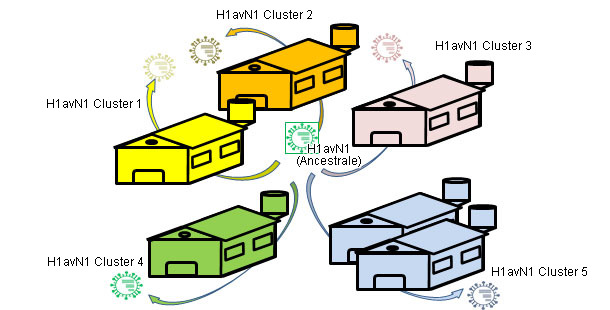
Figura 3. Le infezioni endemiche facilitano l'evoluzione isolata per derivazione in ogni allevamento - eterogenicità genetica ed antigenica correlata con l'allevamento.
In conclusione, i virus dell'influenza suina presentano un'elevata diversità genetica ed antigenica generata da meccanismi evolutivi intrinsechi del virus. Detta diversità è potenziata da fattori epidemiologici come le situazioni di circolazioni endemiche e/o l'isolamento geografico, vedendosi accentuata nel tempo. L'interazione di tutti questi fattori possono avere implicazioni nelle mancanze di riconoscimento da parte del sistema immunitario: ossia in assenza di protezione crociata tra ceppi appartenenti ad uno stesso subtipo.

