

La risposta umorale specifica inizia quando un linfocite B naïve riconosci il suo antigene specifico nei tessuti ed organi linfoidi secondari (vedere "Riconoscimento dei patogeni").
Ogni linfocite B ha un recettore antigenico (BCR: Recettore di cellule B), un'immunoglobulina di superficie (IgM o IgD), che si unisce ai domini specifici dell'antigene specifico chiamati determinanti antigenici o epitopi.

La specificità del sito di unione antigenica dipende dai geni che codificano i domini variabili (VH e VL); nei mammiferi la ricombinazione e/o conversione genetica possono generare un grande numero di geni e, di conseguenza, molti BCRs diversi, capacitando il soggetto a riconoscere quasi tutti gli antigeni esterni.
Dopo il riconoscimento, ogni linfocite B specifico si attiva, realizza una proliferazione clonale e si differenzia in cellula plasmatica (o Cellula Secretrice di Anticorpi, ASC) in grado di sintetizzare e produrre immunoglobuline (anticorpi), specifiche per l'epitope.
Le cellule T aiutano ad ottimizzare la risposta umorale specifica.
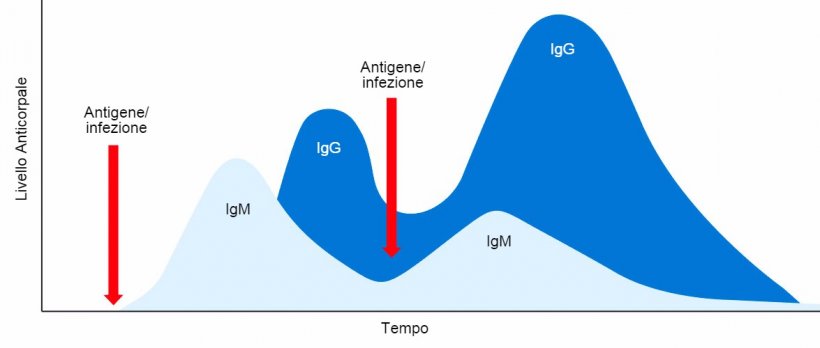
Inizialmente, le cellule B secernono IgM in forma monomerica e con specificità antigenica identica.
In relazione al tipo di secrezione di anticorpi, gli antigeni si classificano come dipendenti o indipendenti dalle cellule T, secondo quanto necessitano, o no, di aiuto delle cellule T per iniziare la risposta con anticorpi.
Per certi antigeni, quelli chiamati indipendenti dalle cellule T (come per esempio i lipopolissacaridi batterici, flageline, polissaccaridi dei pneumoccochi, ecc...) possono solo indurre gli anticorpi IgM con un'affinità modesta e senza cellule B memoria.
Quasi tutti gli antigeni esterni sono dipendenti dalle cellule T e richiedono, per l'attivazione completa, dei linfociti B, un segnale aggiuntivo (citochine) proveniente dai linfociti T collaboratori specifici che riconoscono lo stesso antigene (cooperazione delle cellule T e B).
In quest'ultimo caso, la cellula B agisce come una cellula presentatrice di antigeni, presentando l'antigene elaborato alle cellule T collaboratrici che, quando sono riconosciuti, permette il cambio isotipico dell'Ig (commutazione della classe di anticorpi) da IgM ad immunoglobuline di classe diversa, IgA, IgE o IgG, dipendendo dalla citochina secreta dalla cellula T collaboratrice e coinvolte nella stimolazione delle cellule B (IL-4 o IFNγ rispettivamente). Un'altro segnale basico per il cambiamento dell'anticorpo è l'interazione del CD154 (CD40L) sul recettore CD40 delle cellule B.
Con questo "aiuto", le cellule B proliferatitive provano una ipermutazione somatica dell'immunoglobulina di superficie, per cui passeranno ad essere dotate di un isotipo specifico. Questi linfociti B sono successivamente selezionati in base all'affinità ottimale di Ig con l'antigene al momento del contatto stretto con le cellule dendritiche follicolari che presentano l'antigene. Solamente i linfociti B con un'elevata affinità di anticorpo per l'antigene e che sono in grado di elaborarlo e presentarlo, saranno selezionati positivamente. Pertanto, solo questi ultimi linfociti B sopravviveranno, prolifereranno e si differenzieranno in cellule plasmatiche, sintetizzando e secernendo anticorpi di un'unica classe isotipica, con una specificità unica ed affinità elevata, migliorando la capacità di aderire all'antigene, neutralizzando e distruggendo i patogeni.
Quando termina la risposta immunitaria primaria, alcune di queste cellule B e cellule plasmatiche attivate e selezionate in questo primo contatto, sopravviveranno come cellule memoria di vita lunga, pronte per essere attivate alle successive esposizioni allo stesso antigene (risposta immunitaria secondaria) producendo la stessa classe di anticorpi, con un maggior titolo di affinità.
In conclusione, nella risposta umorale primaria, si produce inizialmente una unica classe di IgM in forma monomerica e con identica specificità antigenica. Quindi, dopo alcuni giorni, grazie alla commutazione isotipica, si producono classi distinte di Ig, per esempio l'IgG. La risposta umorale secondaria indotta da una esposizione precedente allo stesso antigene, sarà caratterizzato principalmente dalla produzione di isotipo di IgG, però con un titolo ed affinità più elevate (figura 1).
Gli anticorpi possono agire nei confronti dei patogeni con differenti meccanismi difensivi:
- Inibizione dell'infettività dei patogeni unendosi a loro (neutralizzazione, blocco dell'adesione, esclusione immunitaria).
- Attivazione della via alternativa (opsonizzazione) o classica del complemento con lisi del patogeno.
- Mentre ricoprono il patogeno (o cellule infette), gli anticorpi permettono che le cellule accessorie, che riconoscono porzioni del frammento cristallizzabile (Fc), ingeriscono e uccidono il patogeno per opsonizzazione e fagocitosi, oppure uccidono le cellule infette tramite ADCC ("Citotossicità Cellulare dipedente da Anticorpi").
- Regolazione della risposta immunitaria attraverso l'interazione con i recettori Fc (FcRs).
Gli FcRs si esprimono in molte cellule effettrice dell'immunità innata, come i mastociti, neutrofili, monociti, macrofagi e cellule dendritiche. Di fatto, quando gli FcγRs (recettori Fc per IgG) si uniscono ai complessi immunari, scatenano l'attivazione delle cellule innate e la loro risposta effettrice (fagocitosi, esplosione ossidativa, citotossicità, liberazione di peptidi vasoattivi e chemioattraenti). Gli FcRs delle cellule dendritiche promuovono la cattura degli antigeni, l'elaborazione e la presentazione alle cellule T citotossiche e collaboratrici.
Al contrario, le FcR inibenti per IgG, espresse nelle cellule B, sono implicate nella regolazione dell'attivazione e sviluppo delle cellule B e nella sopravvivenza delle cellule plasmatiche.


